Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
Nowadays colorimetric assay has been applied
2019-10-29
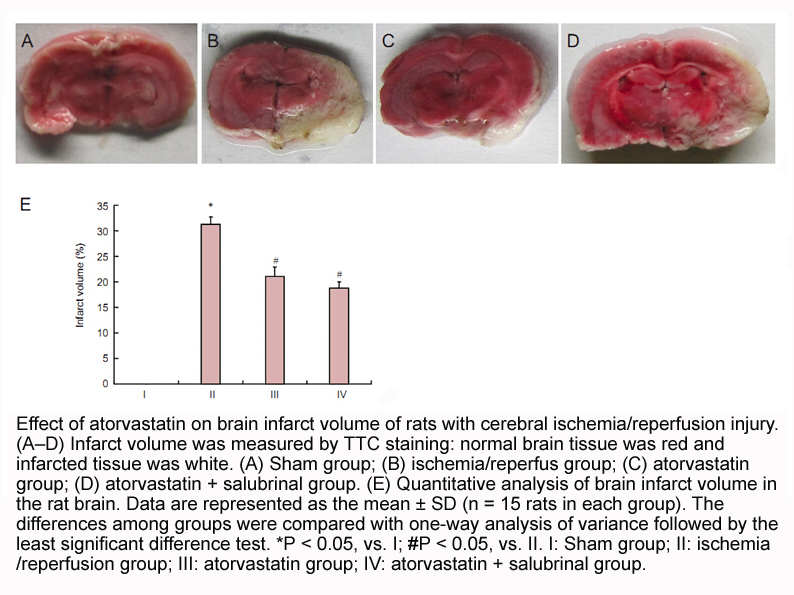
Nowadays, colorimetric assay has been applied in detecting DNA MTase activity. Li et al. used DNA modified gold nanoparticles (AuNPs) coupled with enzyme-linkage reactions to detect the activity of methylases [20]. In a previous study, our group presented a label-free colorimetric method, using unmo
-
Fig D shows that E lagascae VIR E gives
2019-10-29
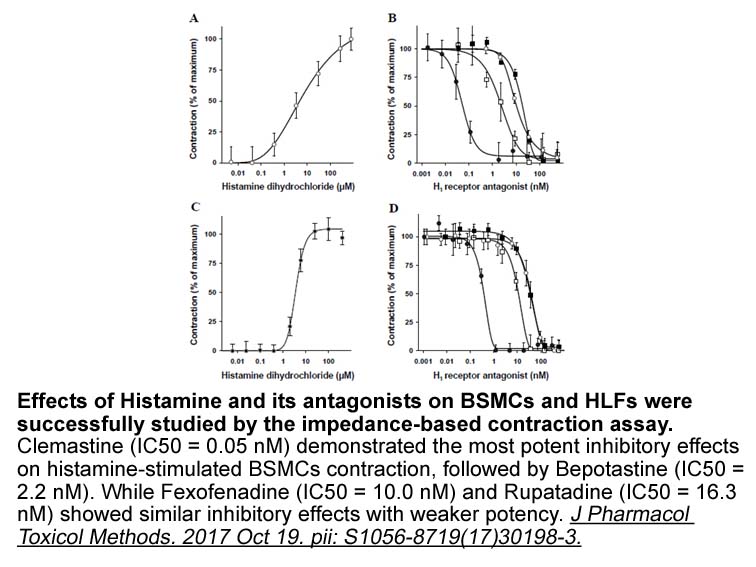
Fig. 2D shows that E. lagascae VIR E gives two species of radioactive TAG: one represents 45% of total labelled TAG and the smaller more-polar spot (15%) is probably derived from the endogenous 1,2-divernolin instead of the exogenous 1,2-diolein. In this case, the total scintillation counts were gre
-
Our conclusion is in disagreement with that of a recent
2019-10-29
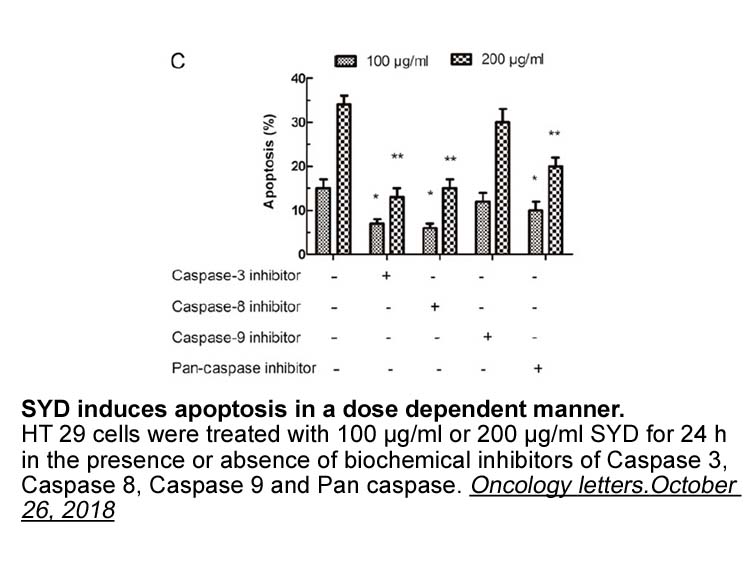
Our conclusion is in disagreement with that of a recent paper by Park et al. [10], which suggests that nitric oxide (NO) inhibition of VHL recruitment occurs by nitrosylation of the 520 cysteine in HIF-1α, and that SNAP does not directly inhibit hydroxylation of HIF peptide by HPH-2. Although this d
-
SKL2001 br Materials and methods br Results br Discussion
2019-10-29
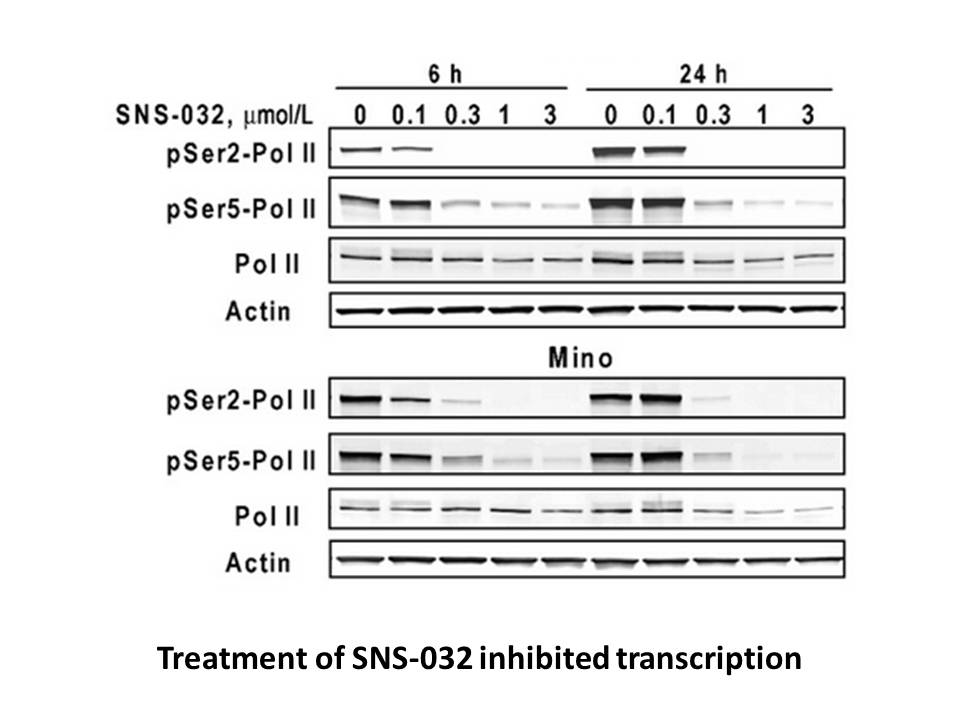
Materials and methods Results Discussion Adiponectin is one of the most studied adipocytokines. Its release from adipocytes is down-regulated in obesity and other adverse metabolic conditions leading to decreased serum concentrations (Turer and Scherer, 2012). Several factors and conditions
-
Cyanine 5-dCTP mg br Conclusion This study demonstrates the
2019-10-29
Conclusion This study demonstrates the presence of functionally viable ET receptors in the surgically extracted tissue from patients with CTEPH. The ET receptor was associated with smooth muscle cells, mainly the contractile phenotype of SMC that surrounds the recanalised Cyanine 5-dCTP mg and c
-
In humans the ER is
2019-10-29
In humans, the ERα is encoded by the gene ESR1, located on chromosome 6, locus 6q25.1 (Gosden, Middleton, & Rout, 1986). In addition to the full-length ERα isoform (66kDa), several shorter isoforms (36kDa, 46kDa) have been identified as a result of the presence of alternate start codons, or as produ
-
br In order to determine the physiological roles
2019-10-29
In order to determine the physiological roles of WT (wild type) EWSR1, Dr. Lee\'s group established a conventional Ewsr1 KO mouse [38]. Since then, the Ewsr1 KO (−/−) mouse model has successfully been utilized to decipher many important in vivo roles of EWSR1 (Fig. 1). EWSR1 deficiency alters m
-
Recent data obtained with etifoxine suggest that facilitatio
2019-10-28
Recent data obtained with etifoxine suggest that facilitation of GABAergic inhibition may be also associated with an indirect mechanism implying activation of the peripheral benzodiazepine receptor with a subsequent increase in neurosteroid production (Schlichter et al., 2000, Verleye et al., 2005).
-
br Methods Adult male B
2019-10-28

Methods Adult male B6, 129 CRHtklee mice (25–30g at the beginning of the experiments) that were wild-type (CRF1R+/+) and recessive homozygous or knockout (CRF1R−/−) were housed six per cage. Mice were 3–5months old and derived from mating CRF1R+/− breeders (Jackson Laboratory, California). Wild-t
-
Parathyroid hormone (1-34) (human) Secondary and primary stu
2019-10-28
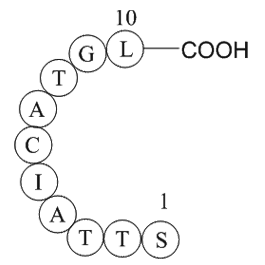
Secondary and primary studies have considered the decision to select the provider and to select components after the component origin is chosen. Examples of secondary and primary studies outside our scope, but still related to decision-making are listed in Tables 1 and 2. The secondary and primary s
-
br Conclusions CK inhibition protects
2019-10-28
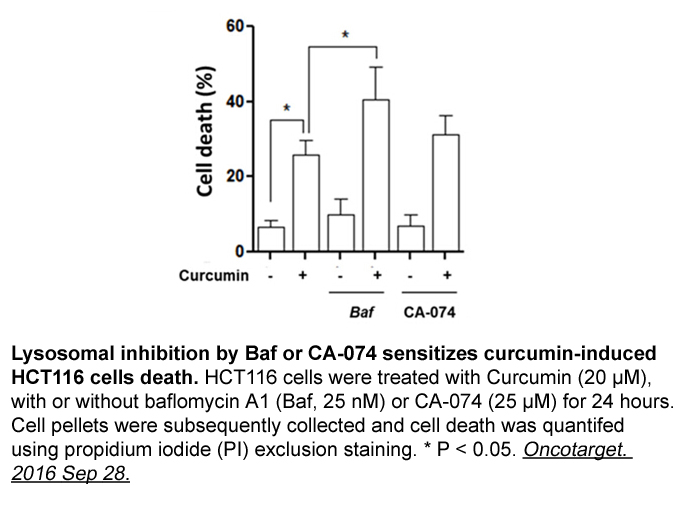
Conclusions CK2 inhibition protects young and aging WM function against an ischemic episode by preserving oligodendrocytes and axonal structure by maintaining mitochondrial integrity. CK2 recruits CDK5 and AKT/GSK3ß signaling to mediate WM ischemic injury in a differential spatiotemporal manner s
-
Two additional hydrogen bonds are
2019-10-28
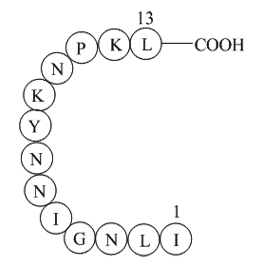
Two additional hydrogen-bonds are formed by the -substituted methoxy-group with Ser and Asp. Ser exerts the role of a gate keeper in CK1: Entry of the ATP in the binding-site causes a conformational change into the closed form by Ser, which locks the binding-site and prevents ATP-diffusion out of th
-
Therefore the relationship between ChEs
2019-10-28
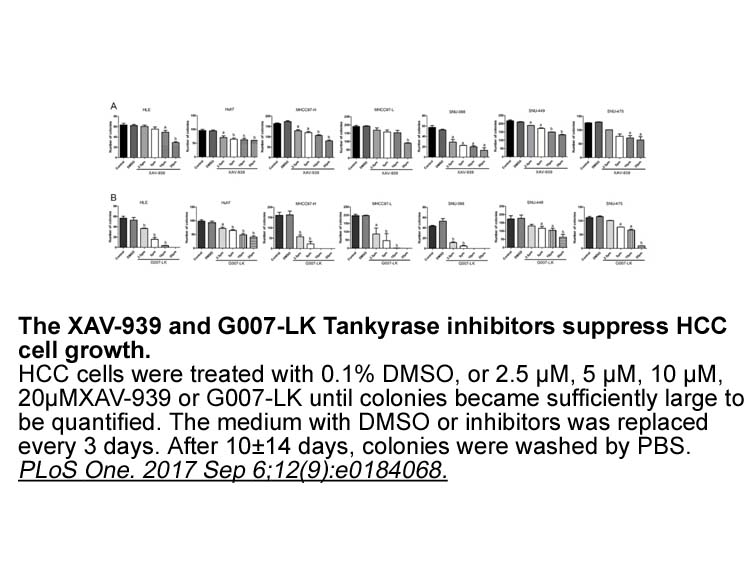
Therefore, the relationship between ChEs activity and polymorphisms in POT1, TERF1, TERT genes were explored in order to screen susceptible biomarkers in this study. Materials and methods Results Discussion However, genetic toxicity induced by OPs has not been fully investigated so far. Ca
-
In conclusion the results obtained in
2019-10-28
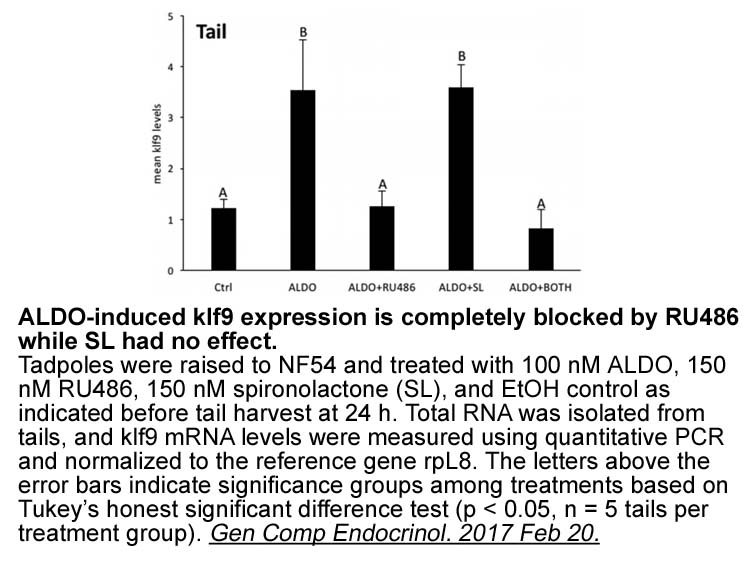
In conclusion, the results obtained in this study and those recently reported by our group (Abdel-Samad et al., 2012) clearly indicate that a dialogue indeed exists between the systems of NPY and ET-1 at the level of human endocardial endothelial cells, since these cells, whether isolated from the r
-
matrix metalloproteinase Astrocytes play a key role in the b
2019-10-28
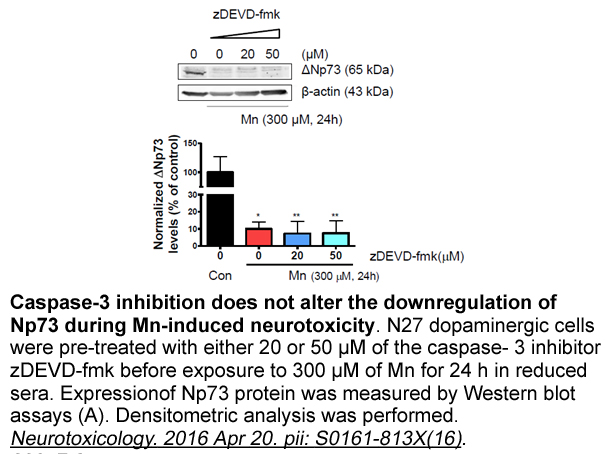
Astrocytes play a key role in the brain, as these cells are involved in fluid, ion, pH, and neurotransmitter homeostasis, synapse function, energy and metabolism and blood-brain barrier (BBB) maintenance (Sofroniew and Vinters, 2010). PA is able to activate different damaging responses in astrocytes
15650 records 940/1044 page Previous Next First page 上5页 936937938939940 下5页 Last page