Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
In pregnancy maternal uterine spiral
2019-08-23
In pregnancy, maternal uterine spiral artery (SA) remodeling is essential for ensuring efficient blood flow to the developing fetus. A subset of CD49a+Eomes+ decidual NK zingerone (dNK) that actively secretes GFs, such as pleiotrophin and osteoglycin, have recently been described in humans and in mi
-
Mulan protein level is regulated
2019-08-23
Mulan protein level is regulated with conditions that stimulate mitophagy such as treatment of SB 415286 with CCCP [12]. We found that exogenously expressed GFP-GABARAP and RFP-Mulan could co-precipitate and co-localize in mammalian cells and the degree of co-localization increased when the cells w
-
The BMP family comprises significant growth factors for the
2019-08-23
The BMP family comprises significant growth factors for the regeneration of bone and cartilage. Among these, BMP- has been known as an early marker during the proliferation of osteoblast cells, and one of the most potent stimulators of osteoblastic cells differentiation [40]. Studies with COX-2 knoc
-
br Transparency document br Main
2019-08-23

Transparency document Main Text The centrosome is a non-membrane-bound organelle found in most animal cells. It has several important functions, including control of cilia formation, microtubule organization and nucleation, spindle assembly and transport of organelles and vesicles. Centrosomes
-
br Material and methods br Results br Discussion Ligase IV
2019-08-23
Material and methods Results Discussion Ligase IV is essential for the final ligation of DSBs through the NHEJ pathway. Its co-factor, XRCC4, has been shown to be important for increasing its activity and stability. Interaction between these two factors have been mapped to the linker region
-
To further determine the cell death
2019-08-23
To further determine the cell death, the Lonidamine in in flasks were harvested after OGD or LTD4 treatment, washed twice with PBS, and evaluated with Annexin V (AV)-FITC apoptosis detection kit I (BD Biosciences Pharmingen, USA) on a flow cytometer (FACSCalibur, Becton–Dickinson, USA). Briefly, 10
-
Taken together our results demonstrate that the changes of
2019-08-23
Taken together, our results demonstrate that the changes of striatal dopamine release and locomotor activity observed following chronic nicotine treatment and consequent acute withdrawal are mediated by CRF1, but not CRF2. Previous studies have already indicated that blocking CRF1 would prevent some
-
br Results and discussion br Conclusion br
2019-08-23
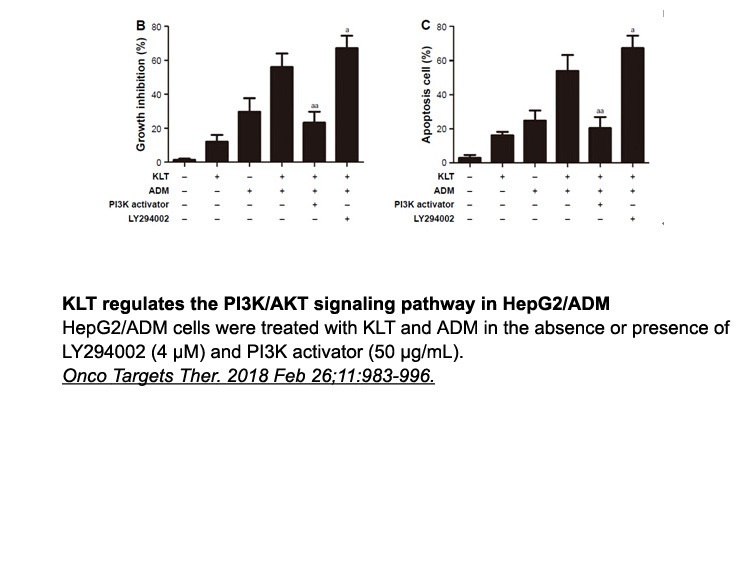
Results and discussion Conclusion Experimental section Pharmacological studies Introduction Survival analysis is a set of statistical methods that aim at modeling the relationship between a set of predictor variables and an outcome variable and, in particular, prediction of the time w
-
It is interesting that of the control groups
2019-08-23
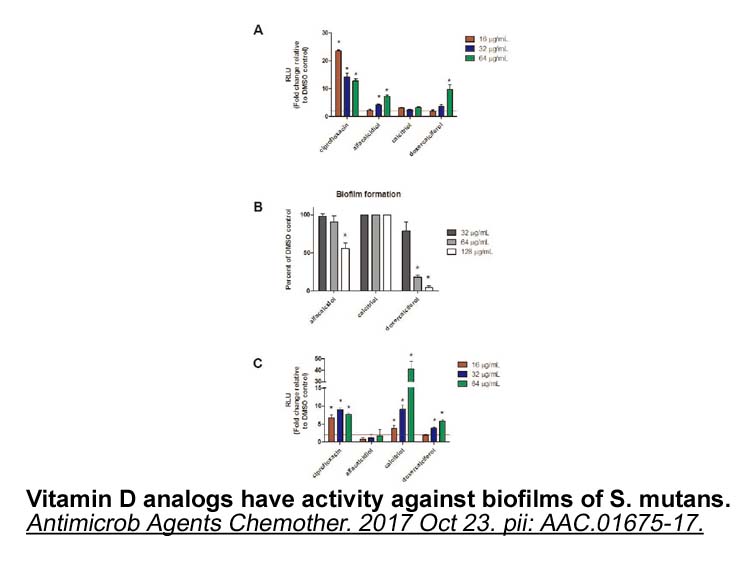
It is interesting that of the control groups, female TG controls only had significantly higher MB-COMT protein than non-TG male controls (Supplemental Table 9, P = 0.005), while these two groups have almost equal expression of S-COMT protein (8.54- vs. 8.34-fold) and their COMT enzyme activity is al
-
br Materials and methods br
2019-08-23
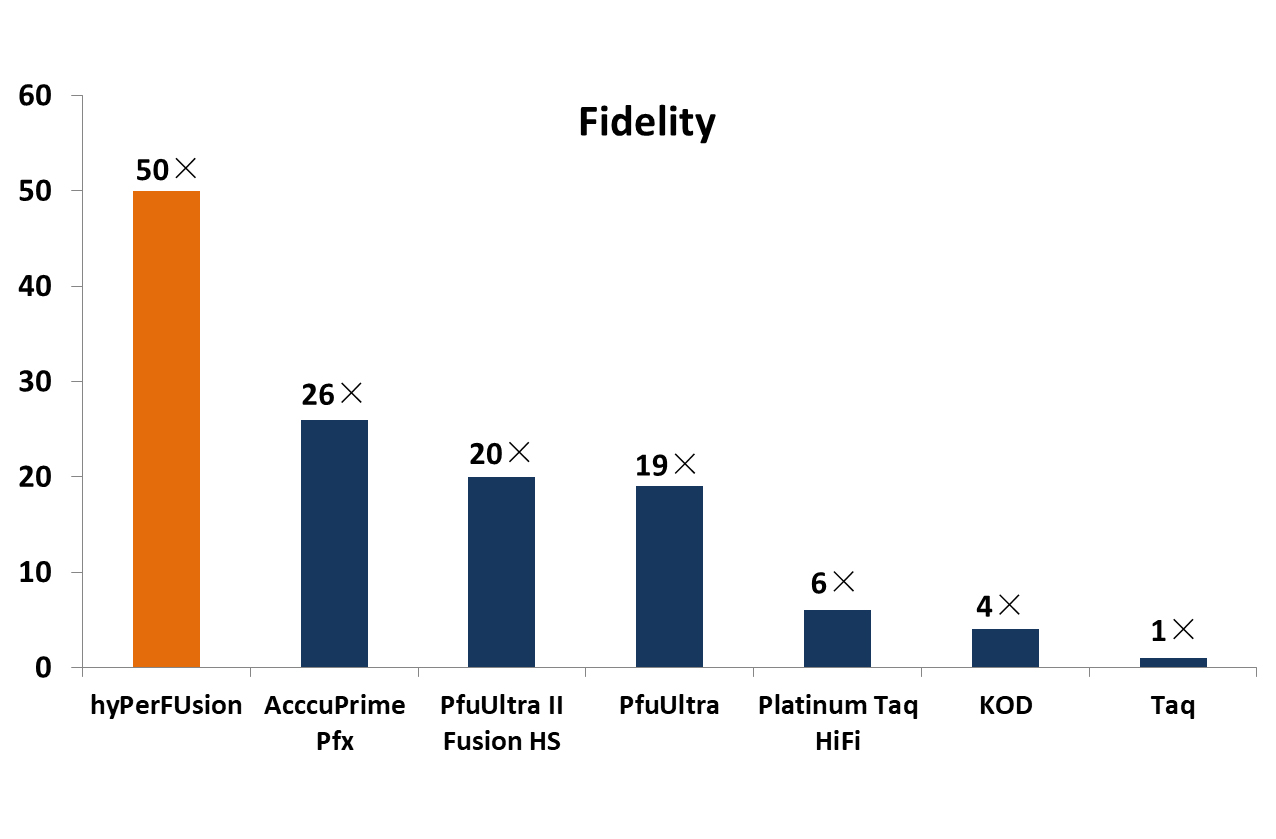
Materials and methods Results Recombinant, N-terminally His6-tagged vCPH was purified to near homogeneity via a modification of the reported procedure and its identity confirmed by intact protein mass spectrometry (Supplementary Fig. 1). As reported, vCPH catalysed two hydroxylations of a PAPK
-
It should be noted that the long lifetime binding responsive
2019-08-23

It should be noted that the long-lifetime binding-responsive luminescence of ARC-1528 and ARC-1530 becomes evident only upon illumination with radiation at near-UV range where the excitation of ATB or ASB moiety is possible. At higher wavelength range (visible light), the fluorophore (5-TAMRA) can b
-
NPC L expression is both tissue and
2019-08-23
NPC1L1 expression is both tissue- and species-specific. Consistent with its function, NPC1L1 is mainly localized to the brush border membrane of enterocytes and the canalicular membrane of hepatocytes. NPC1L1, which is expressed in the liver of humans and rats but not mice, mediates cholesterol Sarp
-
A large fraction of UCEs
2019-08-23
A large fraction of UCEs are transcribed (T-UCEs) in a tissue specific manner, and are deregulated in several human cancers (Calin et al., 2007, Fabbri et al., 2008, Fassan et al., 2014, Olivieri et al., 2016). Indeed, it has been shown that T-UCEs may also act as long non-coding RNAs (lncRNAs) regu
-
Why are Tfh cell dynamics so fundamentally
2019-08-22
Why are Tfh cell dynamics so fundamentally different in naive and immune animals? Initially, GC ITE must pass stringent affinity and specificity checkpoints to ensure only high-affinity non-self-reactive cells are selected. Therefore, restricting primary Tfh cells with the greatest helper capacity
-
The differences between ground and
2019-08-22
The differences between ground and transition state properties are exemplified by a comparison of our human–yeast results with those of a structural study of possible differences in ubiquitin-Uba-1 binding in yeast (known structure) with human (simulated in [12]). Their discussion focuses on Ubiquit
15649 records 978/1044 page Previous Next First page 上5页 976977978979980 下5页 Last page